





Занимаемое популяцией пространство предоставляет ей средство к жизни. Каждая территория может прокормить определенное число особей. Полнота использования ресурсов зависит от размещения особей в пространстве. Оптимальным для популяции является такой интервал между соседними особями, при котором они не влияют отрицателъно друг на друга. Чаще всего члены популяций распределены в пространстве неравномерно. У подвижных животных имеются разнообразные способы упорядочивания распределения в пространстве. Все подвижные животные делятся на две группы - Оседлых И Кочевых. При оседлом существовании животные используют ограниченный участок среды. На хорошо знакомой территории особи спокойно ориентируются, быстро находят еду, быстро прячутся от хищников. Однако при оседлом образе жизни быстрого истощаются ресурсы, если плотность популяции сильно возрастает. Территориальное поведение животных Включает два типа активности: 1) направленную на обеспечение собственного существования (поиск пищи, исследование территории) и 2) на установление отношений с соседними особями. "Закрепляют" участки животные разными способами: 1) охраной границ занимаемой территории и прямой агрессией по отношению к чужаку; 2) особым поведением, демонстрирующим угрозу; 3) системой специальных сигналов и меток, свидетельствующих о занятости территории. Территориальное поведение животных сильно выражено в период размножения животных. Оно обусловлено их физиологическим состоянием и регулируется гормональным путем. У оседлых видов 4 типа пространственного распределения: диффузный, мозаичный, пульсирующий и циклический. В популяциях Диффузного типа Животные распределены в пространстве дисперсно, не образуя обособленных поселений. Он характерен для мелких млекопитающих открытых пространств. Мозаичный тип Размещения возникает тогда, когда пригодные для заселения биотопы распределены в пространстве резко неравномерно. Под воздействием человека формируются мозаичные ареалы у многих видов. Пульсирующий тип Характерен для популяций с резким колебанием численности. При этом может меняться мозаичный тип поселения на диффузный. В период резкого падения численности животные концентрируются на наиболее благоприятных участках, которые получили название "стации переживания". Циклический тип Пространственной структуры характерен для оседлых животных, попеременно используют разные участки территории в течение года. При этом типе использования территории сохраняется баланс между потреблением кормов и его ежегодным восстановлением. При кочевом образе жизни отсутствует зависимость от запаса корма на конкретной территории. Для кочевок животные группируются в стаи и стада. В пасущихся стадах копытных, движущихся косяках рыб и перелетных птиц между особями поддерживается определенное расстояние, при котором они не мешают друг другу, обладая свободой передвижения. Некоторые виды образуют широко кочующие группы (нагульные миграции рыб). Масштабы и длительность миграций зависит от обилия корма и численности стада. При резких нерегулярных изменениях запаса корма наблюдаются нерегулярные перемещения популяции и временное оседание в тех местах, где запасы пищи еще не исчерпаны. Радиус репродуктивной активности – расстояние, преодолеваемое особью для встречи полового партнера.
Экзамен по биологии
93. Динамика популяций во времени. Удельная рождаемость и смертность, скорость изменения численности популяции
Постоянство популяционных функций связано с численность и плотность населения. Это положение предусматривает динамический характер основных популяционных параметров: численность популяции и ее выражение не остается одинаковым во времени. Свойственные популяции процессы постоянного воспроизведения сопровождаются постоянным отмиранием особей. Емкость угодий меняется в сезонном и многолетнем масштабе, что определяет динамику параметров плотности населения. В популяциях постоянно происходят процессы притока особей извне и выселения определенной части населения за пределы популяции. Все эти процессы придают динамический характер популяции. Особи в популяции отличаются друг от друга по полу, возрасту, генетическим особенностям и месту в функциональной структуре популяции. На этой основе формируются типы воспроизводства и смертности, масштабы и формы пространственной дисперсии, общий уровень биологической активности и др. К основным динамическим характеристикам популяции относятся рождаемость и смертность. Рождаемость – это способность популяции к увеличению численности. В популяционной экологии это появление на свет новых особей любого организма независимо от того, рождаются ли они, вылупляются из яиц, прорастают из семян или появляются в результате деления. Максимальная рождаемость — это теоретический максимум скорости образования новых особей в идеальных условиях. Максимальная рождаемость постоянна для данной популяции. Экологическая рождаемость обозначает увеличение численности популяции при фактических или специфических условиях среды. Эта величина не постоянна и варьирует в зависимости от размерного и возрастного состава популяции и физических условий среды. Рождаемость выражают как скорость, определяемую путем деления общего числа появившихся особей на время (Абсолютная рождаемость), либо как число появившихся особей в единицу времени на 1 особь в популяции (Удельная рождаемость). Различие между абсолютной и удельной рождаемостью легко проиллюстрировать на таком примере: в городе с населением 10 000 появилось 400 новорожденных. Абсолютная рождаемость составит 400 в год, а удельная — 0,04 (4 на 100, или 4%). Различие между максимальной и реализованной рождаемостью можно проиллюстрировать при изучении природной популяции птицы на гнездовьях городского парка, и популяции мучного хрущака. Птицы отложили 510 яиц, оперилось 265 птенцов, поэтому экологическая рождаемость составляет 52%. Жуками было отложено 12 000 яиц, из которых вылупились только 773 (или 6 %) личинки. Для видов, которые не охраняют яйца и не заботятся о потомстве, характерна высокая потенциальная и низкая реализованная рождаемость. Смертность отражает гибель особей в популяции. Смертность можно выразить числом особей, погибших за данный период или в виде удельной смертности для всей популяции. Экологическая смертность — гибель особей в данных условиях среды. Эта величина не постоянна и изменяется в зависимости от условий среды и состояния самой популяции. Теоретическая Минимальная смертность — величина, постоянная для популяции; она представляет собой гибель особей в идеальных условиях, при которых популяция не подвергается лимитирующим воздействиям. Даже в самых лучших условиях особи будут умирать от старости. Этот возраст определяется Физиологической продолжительностью жизни, Которая намного превышает среднюю Экологическую продолжительность жизни.
Рост численности популяции можно представить как постоянно идущий процесс, масштабы которого зависят от свойственной данному виду скорости размножения, которая определяется как удельный прирост численности за единицу времени: r=dN/Ndt: где r — «мгновенная» удельная скорость роста популяции, N — численность ее и t - временной промежуток, в течение которого учитывалось изменение численности. При условиях неограниченного роста изменение численности популяции во времени выражается экспоненциальной кривой, Описываемой уравнением Nt = N0*реRt, Где No—исходная численность, Nt — численность в момент времени T, E — основание натуральных логарифмов. ЭКспоненциальная модель роста популяции Отражает ее потенциальные возможности размножения. Показатель мгновенной удельной скорости роста популяции r Определяют как Репродуктивный потенциал популяции Или ее Биотический потенциал. Экспоненциальный рост популяции возможен при условии неизменного значения коэффициента r.

94. Понятие сообщества. Видовое богатство. Индексы разнообразия
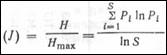
Сосуществующие организмы одного вида характеризуются плотностью, соотношением полов, возрастной структурой, рождаемостью, смертностью, иммиграцией и эмиграцией. Взаимодействия на популяционном уровне влияют на высокий уровень организации — Сообщество - совокупность популяций разных видов, сосуществующих в пространстве и времени. Сообщества изучает синэкология. Сообщество — сумма образующих его видов и совокупность взаимодействий между ними, т. е. оно имеет Эмерджентные Свойства. Примерами эмерджентных свойств будут видовое разнообразие, пределы сходства конкурирующих видов, структура пищевой сети, биомасса и продуктивность сообщества. Экосистема - сообщество вместе с его физической средой обитания. В пределах иерархии местообитаний можно выделить сообщество любого размера, масштаба или уровня. Климат - основной фактор, определяющим границы распространения разных типов растительности. При изучении сообществ все организмы, обитающие в каком-либо районе, рассматриваются вместе. Исследование сообщества ограничено одной систематической группой в его составе (птицами, насекомыми или растениями) или группой с особым типом жизнедеятельности (травоядными, детритофагами). Видовое богатство сообщества и индексы разнообразия. Один из способов охарактеризовать сообщество — пересчитать входящие в него виды или составить их список, что позволяет описывать и сравнивать сообщества по их видовому богатству. Число отмеченных видов зависит от числа взятых проб или размеров изученного местообитания. Обычные виды должны встретиться в первых нескольких пробах; чем больше проб, тем более редкие таксоны попадут в список. Кривая видового богатства выходит на плато. Когда состав сообщества характеризуется числом входящих в него видов, полностью игнорируются количественные отношения между ними. При этом теряется информация о редкости одних видов и обычности других. Лучше описать сообщество с точки зрения биомассы или продукции разных видов на единицу площади. Параметр сообщества, учитывающий число видов и соотношение их обилия, — индекс разнообразия Симпсона. Его рассчитыпают, определяя для каждого вида долю его особей или биомассы в общей численности или биомассе выборки. Если доля i-го вида — РI,То
Индекс разнообразия Симпсона (D):
![]()
Где S—общее число видов в сообществе (т. е. видовое богатство). Величина этого индекса зависит и от видового богатства и от равномерности в соотношении обилий разных видов. При постоянном числе видов D возрастает с увеличением выровненности в количественном соотношении разных видов, а при постоянной равномерности — с ростом видового богатства. Равномерность распределения (выровненность) можно количественно оценить при помощи индекса Симпсона — доля максимально возможной величины D, Достигаемой при одинаковой численности всех видов. Если Dmaх=S, то равномерность распределения (Е) (этот показатель принимает значения от 0 до 1)
![]() .
.
ругой индекс разнообразия— индекс Шеннона (Н), Зависит от совокупности значений РI:
![]() Разнообразие
Разнообразие
В этом случае равномерность распределения (J):
32. Характеристика отрядов насекомоядные и рукокрылые
НАСЕКОМОЯДНЫЕ ЗВЕРИ (Insectivora), отряд млекопитающих; включает 7–8 семейств, в том числе: щелезубы, тенреки, ежи, землеройки, кроты, выхухоли, всего около 300 видов. Это самые древние и примитивные из плацентарных млекопитающих. Длина тела насекомоядных от 3 до 45 см. У многих представителей 44 зуба. Тело большинства зверьков покрыто густым бархатистым мехом, у некоторых — жесткими щетиновидными волосами и короткими иглами. Для многих характерны специфические (мускусные и пахучие) железы. Головной мозг с небольшим обонятельным отделом, размеры полушарий невелики. Из органов чувств наиболее развиты органы обоняния и осязания. Органы зрения почти у всех сформированы слабо. Насекомоядные распространены в Африке, Евразии, Северной Америке и севере Южной Америки, отсутствуют в Австралии и почти во всей Южной Америке. Восемь видов занесены в Международную Красную книгу.
РУКОКРЫЛЫЕ (Chiroptera) — отряд млекопитающих; включает около 850 видов, которые делятся на два подотряда — крыланы и летучие мыши. К рукокрылым относятся мелких и средних размеров зверьки, передние конечности которых превращены в крылья. Рукокрылые способны к полету; между плечом, предплечьем, пальцами, боками тела и задними конечностями натянута тонкая летательная перепонка. Ушные раковины большие, у многих с хорошо развитым кожным выступом — козелком. Хвост у большинства видов длинный. Череп с крупной мозговой коробкой. Глаза у плотоядных видов большие и зрение умеренно развито. У большинства видов глаза маленькие. В пространстве ориентируются с помощью ультразвуковой эхолокации (кроме крыланов). Рукокрылые распространены на всех материках (кроме Антарктиды) и почти на всех крупных островах к северу от лесотундровой зоны. Они активны в сумерки и ночью. Днем большинство видов находится в убежищах: пещерах, дуплах деревьев и др. Здесь же впадают в зимнюю спячку. Питание очень разнообразно. Часть видов предпочитает растения и тропические плоды (листоносы), насекомых (нетопыри, вечерницы), вампиры питаются кровью млекопитающих. Стадность (образование колоний) характерна для большинства видов. Размножение у многих рукокрылых — обитателей тропических стран происходит 2 раза, у остальных видов — 1 раз. В каждом приплоде родится по одному детенышу (редко 2). У большинства видов детеныш рождается крупным и быстро растет. Врагов у рукокрылых немного (совы, сычи). Большинство видов являются полезными. Нетопыри уничтожают вредных насекомых, листоносы, поедая плоды диких деревьев, распространяют древесные породы и др. Вредными считаются вампиры. Помет рукокрылых — высококачественное удобрение.
95. Пространственная структура сообществ. Градиентный анализ
Бывают сообщества с резкими границами, которые не могут пересечь виды соседних сообществ. Н-р: разделение наземной и водной сред. На суше резкие границы наблюдаются между растительностью сопредельных выходов кислых и щелочных пород, а также в местах соприкосновения серпентиновых (богатых силикатом магния) и несерпентиновых субстратов. Однако в таких ситуациях минеральные вещества попадают через границы, которые становятся более размытыми. В горах Грейт-Смоки-Маунтинс основной облик сообществ определяется древесными породами. На склоне условия для роста растений неоднородны, на распределение пород деревьев сильно влияют высота над уровнем моря и влажность. Растительность различных участков суши неодинакова. Чем круче градиент ключевого фактора среды, тем более четкими окажутся границы распределения различных групп видов. В экстремальных условиях градиенты бывают особенно крутыми. Доминирующие виды своим распределением четко обозначают границы сообществ. Когда ключевой фактор среды меняется по градиенту, то должны существовать точки, в которых происходит коренное изменение относительной конкурентоспособности видов, в результате чего один доминант сменяется другим и само взаимодействие видов приведет к образованию и закреплению границы между зонами их распределения. Н-р: сообщество многолетних злаков может создать между двумя доминантами зоны напряженности. Если внезапные смены одного типа растительности другим существуют, то межсезонные колебания условий всегда будут вызывать непрерывное смещение точки скачка.
96. Изменение сообществ во времени (сукцессия). Основные типы сукцессий
Вид присутствует в сообществе только в том случае, если: 1) он способен достичь данного места; 2) условия и ресурсы этого места для него подходят; 3) он выдерживает конкуренцию и выедание со стороны других видов. Необходимо, чтобы условия, ресурсы и влияние других видов изменялись во времени. Обилие многих организмов в сообществе меняется в течение года по мере того, как особи завершают свои жизненные циклы при переходе от одного сезона к другому. СУкцессия - Несезонная, направленная и непрерывная последовательность появления и исчезновения популяций разных видов в неком местообитании. Деградационная сукцессия. Она происходит быстро —в течение нескольких месяцев или лет. Н-р, мертвая органика (погибшее животное или растение) используется микроорганизмами или животными-детритофагами. Разные виды редуцентов появляются и исчезают поочередно, по мере истощения в ходе разложения органического вещества одних ресурсов и появления других веществ. В этих последовательных сменах участвуют гетеротрофы, то эту сукцессию называют Гетеротрофной. Деградационная сукцессия наблюдается при использовании разными видами грибов опавших хвоинок сосны. Аллогенная сукцессия. Новое местообитание представлено участком субстрата, открытым для заселения зелеными растениями (Автотропной сукцессии) Или другими сидячими организмами. В этих случаях новое местообитание не исчезает, а заселяется. Необходимо отличать автогенные сукцессии, являющиеся результатом биологических процессов, меняющих условия и ресурсы, от Аллогенных, Обусловленных внешними геофизико-химическими обстоятельствами. Н-р, переход от соленых болот (маршей) к лесу. Эстуарии рек быстро заиляются. Происходит осадконакопление на отмелях. В результате соленый марш сдвигается в сторону моря. Пойменный лес перемещается за болотом.
Ближе всего к уровню моря располагаются пионеры заселения солоноватоводных илистых отмелей — камыш и полевица. Они встречаются по всему маршу, доходя до границы произрастания дуба. Анализ остатков растительности в лесной почве доказывает процесс сукцессии. Эта сукцессия определяется внешним абиотическим фактором — осадконакоплением. Автогенная сукцессия. Сукцессии на заново осваиваемых территориях в отсутствие постепенного изменения абиотических факторов называются Автогенными. Если заселяемый участок ранее подвергался воздействию какого-либо сообщества - это Первичная сукцессии (на недавно сформировавшихся песчаных дюнах, застывшей лаве, при отступлении ледника). Когда растительность на местности частично или полностью уничтожена, но хорошо развитая почва, семена и споры остаются, последующая смена видового состава - это Вторичная сукцессия, н-р, уничтожение леса болезнями, ураганом, пожаром или вырубкой. Сукцессии на заново осваиваемых территориях обычно идут несколько веков. Часто стадии сукцессии представлены градиентом сообществ в пространстве (переход от соленого марша к лесу). Первые виды в сукцессии могут настолько изменить условия или доступность ресурсов в местообитаниях, что становится возможным поселение новых видов (процесс Облегчения). Он важен при первичной сукцессии, когда исходные условия суровы, например на обнаженной почве после отступления ледника. Сукцессия на ледниковых отложениях идет следующим образом: первыми поселяются мхи и отдельные растения с поверхностной корневой системой. Затем появляются ивы, сначала стелющиеся, позже — кустарниковые виды. Вскоре в сукцессию вовлекается ольха с тополями. Ольха вытесняется ситхинской
Елью, и в итоге образуется смешанный лес с тсугами. Одна из основных движущих сил сукцессии — изменение почвы первыми колонистами. Ольха имеют симбионты, фиксирующие атмосферный азот, что приводит к накоплению в почве больших его запасов. Ситхинская ель, используя накопленный азот, может поселяться и замещать ольху. Жизненный цикл раннесукцессионных растений короток. Их непрерывное существование зависит от переноса в другие нарушенные места. Неспособные выдержать конкуренцию с появляющимися позднее видами, они вынуждены быстро расти (основное их свойство), эффективно используя доступные ресурсы. Скорость фотосинтеза на единицу площади листа и относительная скорость роста с течением сукцессии в снижаются. Один из факторов успеха позднесукцессионных видов — их теневыносливость. Организмы, появляющиеся первыми, способны истощить ресурсы и заставить голодать появляющихся позднее. Однако «проигравшие» конкуренты становятся «победителями», если особенности их жизненного цикла позволяют им расти в период прекращения активности соперников. Деревья последних стадий залежной сукцессии могут быть разделены на ранне - и позднесукцессионные. Для многих деревьев первого класса характерна многоярусность кроны. Виды (можжевельнику) способны использовать обилие света на ранних стадиях сукцессии древесных пород. Клен сахарный и бук крупнолистный - одноярусные породы. У них листья кроны расположены в один слой, окружающий ствол наподобие шлема. Они успешнее существуют в густом пологе на поздних стадиях сукцессии. Если один и тот же участок одновременно заселяют многоярусные и одноярусные деревья, первые растут быстрее и доминируют до тех пор, пока их не затенят соседи. У раннесукцессионных деревьев расселение семян происходит более эффективно, что повышает вероятность быстрого занятия ими свободного пространства. Они скороспелы. У позднесукцессионных деревьев семена крупнее и менее эффективно расселяются; созревают эти растения позднее.
97. Концепция экосистемы. Ее компоненты. Принципы функционирования
Живые организмы и их неживое (абиотическое) окружение неразделимо связаны друг с другом и находятся в постоянном взаимодействии. Любая единица, включающая все совместно функционирующие организмы на данном участке и взаимодействующая с физической средой таким образом, что поток энергии создает определенные биотические структуры и круговорот веществ между живой в неживой частями, представляют собой Экосистему. Экосистемы представляют собой открытые системы, они должны получать и отдавать энергию, поэтому важной составной частью концепции являются Среда на выходе И Среда на входе. Термин экосистема был предложен Тенсли. Экосистемы, входящие в биосферу, в разной степени открыты для потоков веществ, для иммиграции и эмиграции организмов. Функционирующая экосистема должна иметь вход и пути оттока переработанной энергии и веществ. Масштабы изменений среды на входе и на выходе сильно варьируют и зависят от: 1) размеров системы (чем она больше, тем меньше зависит от внешних частей); 2) интенсивности обмена (чем он интенсивнее, тем больше приток и отток); 3) сбалансированности автотрофных и гетеротрофных процессов (чем сильнее нарушено это равновесие, тем больше должен быть приток извне для его восстановления); и 4) стадии и степени развития системы (молодые системы отличаются от зрелых). Структура экосистемы. С точки зрения Трофической структуры экосистему можно разделить на два яруса: 1) верхний Автотрофный ярус, Или «зеленый пояс», включающий растения, содержащие хлорофилл, где преобладают фиксация энергии света, использование простых неорганических соединений и накопление сложных органических соединений, и 2) нижний Гетеротрофный ярус, Или «коричневый пояс» почв и осадков, в котором преобладают использование, трансформация и разложение сложных соединений. В составе экосистемы можно выделять компоненты: 1) Неорганические вещества (С, N, СО2, Н2О), включающиеся в круговороты; 2) Органические соединения (белки, углеводы, липиды, гумусовые вещества), связывающие биотическую и абиотическую части; 3) Воздушную, водную И Субстратную среду, Включающую Климатический режим; 4) Продуцентов, Автотрофных организмов; 5) Макроконсументов, Или Фаготрофов — гетеротрофных организмов, (животные), питающихся другими организмами или частицами органического вещества; 6) Микроконсументы, сапротрофов, Деструкторов, Или Осмотрофов — гетеротрофных организмов (бактерии и грибы), получающих энергию путем разложения мертвых тканей, либо путем поглощения растворенного органического вещества. Энергетическая классификация экосистем. Источник и качество доступной энергии определяют видовой состав и численность организмов, характер функциональных процессов, протекающих в экосистеме, и процессов ее развития, и образ жизни человека. Выделяют четыре главных типа экосистемы: 1. Природные, движимые Солнцем, несубсидируемые. 2. Природные, движимые Солнцем, субсидируемые другими естественными источниками. 3. Движимые Солнцем и субсидируемые человеком. 4. Индустриально-городские, движимые топливом (ископаемым, другим органическим или ядерным).
98. Биомы. Их определение. Основные виды
БИОМ - совокупность различных групп организмов и среды их обитания в определенных природных зонах и поясах. Ключевая характеристика, позволяющая разграничивать и узнавать наземные биомы, — Жизненная форма Растительности климатического климакса (травы, кустарники, листопадные деревья, хвойные деревья). Климаксная растительность степного биома (злаковника) — злаки. Основа классификации — растительность климатического климакса, однако биом включает в себя эдафические климаксы, и стадии развития, в которых доминируют иные жизненные формы. Например, степные сообщества представляют собой стадии развития лесного биома, а прибрежные леса — составная часть степного биома. Подвижные животные связывают между собой различные ярусы и стадии развития растительности. Птицы, млекопитающие, рептилии и многие насекомые свободно передвигаются между подсистемами и между развивающимися и зрелыми стадиями растительности, а перелетные птицы в определенные сезоны — между биомами разных континентов. Для наземных биомов характерны крупные растительноядные млекопитающие — олени, антилопы, бизоны, домашний скот. По составу флоры и фауны разделяют мир на пять или шесть главных областей, которые приблизительно соответствуют основным континентам. Это тундры, северные хвойные леса, листопадные леса умеренной зоны, степи умеренной зоны, тропические степи и саванны, чапараль и жестколистные леса, пустыни и тропические лесса. Тундры и северные хвойные леса. Между лесами на юге и шапкой полярных льдов на севере расположена полоса безлесной местности. Небольшие районы расположены в высоких горах выше границы леса. Такие районы называются альпийскими тундрами. В Северной Америке и Евразии граница между тундрой и лесотундрой сдвинута к северу на западе. Основными лимитирующими факторами служат низкие температуры и короткий сезон вегетации; осадков тоже может быть мало, но они не относятся к лимитирующим факторам. Грунт на протяжении всего лета остается промерзшим. Биомы северных хвойных лесов. Широким поясом вытянулись через всю Северную Америку и Евразию области северных вечнозеленых лесов. Доминирующая жизненная форма представлена здесь хвойными вечнозелеными деревьями: елью, пихтой и сосной. На протяжении всего года в таких лесах царит густая тень, поэтому кустарниковый и травяной ярусы обычно развиты слабо. Для этого биома характерен высокий уровень годовой продукции. Листопадные леса умеренной зоны. Сообщества листопадных лесов занимают области с большим количеством равномерно распределенных осадков (750—1500 мм) и умеренной температурой, для которой характерны сезонные колебания. Умеренные листопадные леса покрывали весь восток Северной Америки, всю Европу, часть Японии, Австралии и юг Южной Америки. Биомы листопадных лесов изолированы друг от друга, чем тундра и северные хвойные леса. Травянистый и кустарниковый ярусы, почвенная биота развиты хорошо. Многие растения дают сочные мясистые плоды и орехи, желуди или буковые орешки. Животное население первоначальных лесов Северной Америки представлено виргинским оленем, медведем, серой и черной белкой, серой лисицей, рыжей рысью и дикой индейкой. Степи умеренной зоны расположены там, где выпадает промежуточное между пустынями и лесами количество осадков (250— 750 мм); существование степей в этих условиях зависит от температуры, сезонного распределения осадков и влагоемкости почвы. Ключевым фактором здесь является влажность почвы. Обширные степные пространства занимают внутренние части Евразийского и Североамериканского континентов, юг Южной Америки (аргентинские пампасы) и Австралию. В Северной Америке биом степей подразделяется на следующие зоны: высокотравные, смешанные, низкотравные и кустисто-злаковые прерии. Тропические степи и саванны. Тропические саванны (степи с редкими деревьями или группами деревьев) расположены в теплых областях, где в год выпадает большое количество осадков (1000—1500 мм), но имеется один или два продолжительных сухих сезона, когда возникают пожары. Самая обширная область этого типа находится в Центральной и Восточной Африке. Растительность состоит из небольшого числа видов. В покрове доминируют знаки. Ландшафт африканской саванны усеян акациями, бобовыми, баобабами, Молочаями и пальмами. Здесь пасутся многочисленные виды антилоп (гну), зебры и жирафы, львы и другие хищники. Насекомые обильны во время влажного сезона, когда гнездится большинство птиц; рептилии активнее во время сухого сезона. Сезонность определяется дождями, а не температурой, как в степях умеренной зоны. Чапараль и жестколистные леса. В областях с мягким умеренным климатом, где обильны зимние дожди, но лето сухое, растительность состоит из деревьев или кустарников с жесткими толстыми вечнозелеными листьями. Пустыни встречаются в тех областях, где в год выпадает меньше 250 мм осадков. Скудость осадков может быть вызвана: 1) высоким субтропическим давлением (Сахара и австралийские пустыни); 2) географическим положением в области «дождевой тени» (пустыни на западе Северной Америки); 3) большой высотой местности (пустыни Тибета и Боливии, Гоби) . В большинстве пустынь в течение года выпадает какое-то количество дождей, и имеется хотя бы редкая растительность. Влажные тропические леса. Широколиственные вечнозеленые дождевые тропические леса расположены на малых высотах полосой вдоль экватора. Дождей выпадает от 2000 до 2250 мм в год; в течение года отмечается один или несколько «сухих» сезонов (120—130 мм в месяц). Дождевые леса встречаются в трех областях: 1) бассейны Амазонки и Ориноко в Южной Америке и Центральноамериканский перешеек; 2) бассейны Конго, Нигера и Замбези в Центральной и Западной Африке и Мадагаскар; 3) области Индо-Малайская и Борнео—Новая Гвинея. Сезонная периодичность размножения растений и животных связана с колебаниями количества осадков. Дождевой лес характеризуется сильно выраженной ярусностью. Деревья формируют три яруса: 1) редкие очень высокие деревья, которые возвышаются над общим уровнем полога; 2) полог, образующий вечнозеленый покров на высоте 25—35 м; 3) нижний ярус, который становится густым только там, где имеются просветы в пологе.
99. Генетика онтогенеза
Дифференциальная активность генов в ходе онтогенеза. В результате дифференциальной активности генов формируются различные дифференцированные клеточные линии, а на их основе — ткани и органы. Дифференцировкой называют комплекс изменений, вовлеченных в прогрессивные расхождения в структуре и функциях клеток организма. При этом в каждой линии клеток дифференцировка приводит к постоянному сужению спектра транскрипции. Организм взрослых гермафродитов нематод состоит из 959 соматических клеток, у самцов 1031 клетка. Число зародышевых клеток у обоих полов варьирует. Число соматических клеток фиксировано. Каждая клеточная линия (ткань, орган) характеризуется определенным набором генов, активируемых в процессе ее развития. Число генов, вовлеченных в развитие и функционирование органов и тканей человека: мозг – 3195, глаз – 547, почка 712, эмбрион – 1989. ТОТИПОТЕНТНОСТЬ способность отдельных клеток в процессе реализации заключенной в них генетической информации не только к дифференцировке, но и к развитию в целый организм. Тотипотентны оплодотворенные яйцеклетки растений и животных. Для соматических клеток животных характерна тканевая специфичность с ранних стадий эмбрионального развития, и поэтому они не обладают тотипотентностью. Однако стволовые клетки в обновляющихся тканях животных в пределах одного типа ткани могут развиваться в разных направлениях. Например, стволовые клетки кроветворной ткани млекопитающих дают начало эритроцитам и лейкоцитам. Соматические клетки растений способны полностью реализовать свой потенциал развития с образованием целого организма. Специализированные клетки самых разных органов (листа, корня, цветка) способны к размножению в искусственной среде вне организма. При создании оптимального соотношения фитогормонов в питательной среде культивируемые клетки могут образовывать побеги или превращаться в результате соматического эмбриогенеза в зародышеподобные структуры, которые затем развиваются в целый организм. Способность соматических клеток растений проявлять тотипотентность зависит от генотипа. Тотипотентность соматических клеток лежит в основе их использования в генетической и клеточной инженерии. Гомеозисные мутации у дрозофилы. После завершения формирования сегментации, вступают в действие гомеозисные гены — большой класс генов, которые контролируют развитие какой-то части тела из определенного сегмента. В результате гомеозисной мутации из данного сегмента развивается какая-то другая часть тела. Среди гомеозисных генов наиболее известны Bithorax-Complex (BX-C) И Antennapedia-Complex (Ant-C). У дрозофилы личинки и имаго имеют ярко выраженные сегменты: один головной, три грудных и восемь брюшных. Каждый сегмент имаго содержит набор дифференцированных морфологических структур. Мезоторакальный сегмент несет пару крыльев и пару ног, метаторакальный — пару ног и пару гальтеров — особых булавовидных образований, помогающих удерживать равновесие в полете. Есть группа генов, отвечающих за формирование гальтеров и брюшных сегментов. Одним из генов, влияющих на эти процессы, является ВХ-С. Без этого гена эмбрион развивается до определенной стадии и затем гибнет. Если бы этот организм остался жить, то он бы имела 10 пар крыльев и 10 пар ног. Функция гена ВХ-С Заключается в инактивации генов, формирующих ноги и крылья во всех последующих после второго торакального сегментах. Комплекс ВХ-С Содержит три различных гена: Ubx, Abd-A И Abd-B. Каждый из них контролирует формирование определенной группы сегментов. Мутации этих генов заставляют все последующие сегменты формироваться подобно одному из предыдущих. Если все три гена удалены, Нормально развиваются только первый торакальный (Т1) и девятый брюшной (А9) сегменты, контролируемые другими генами, все остальные сегменты (ТЗ и все брюшные) развиваются как Т2. Если ген Ubx Сохраняется, но повреждаются Abd-A И Abd-B, Нормально развиваются все грудные сегменты, а все брюшные представлены самым первым — А1. При повреждении гена Abd-B Нормально развиваются все грудные сегменты, затем брюшные Al, A2 и A3, а все остальные сформированы как сегмент А4.